Tag: DNA taxonomy

It seems that not only are the North American Aneura sharpii, the east Asian Aneura pellioides and the south-east Asian Aneura maxima all distinct species, but there are also two Pellia-like Aneura species in Europe.

The simple thalloid liverwort Aneura has become a flagship genus for DNA barcoding at RBGE. Only a single widespread species, Aneura pinguis, is traditionally recognized in the UK,…

Geography, and particularly climate, have distinguished the extreme western parts of Scotland from the rest of the country for thousands of years. Many of our rarest plant species…
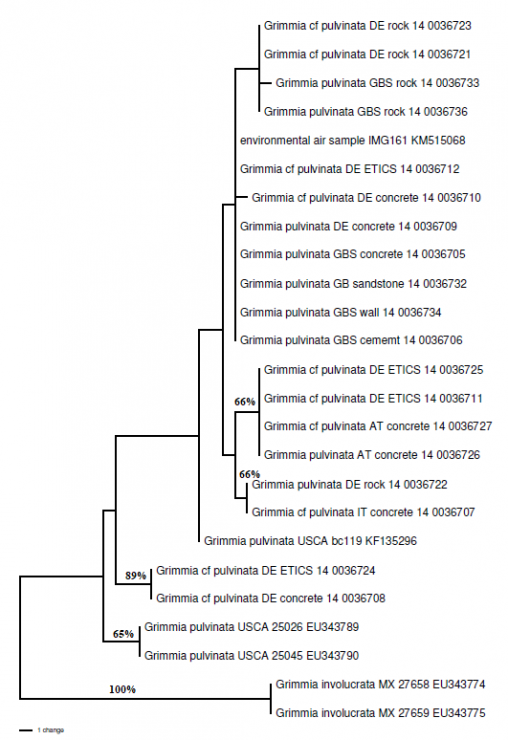
Even with a plant as common, and as commonly overlooked, as this pollution-tolerant urban bryophyte, there is still genetic diversity to explore and explain.

After RBGE’s initial involvement in land plant DNA barcode marker selection, culminating in a couple of 2009 papers that both utilized bryophyte barcoding data sets, we started a…

Enigmatic and isolated although it is, it seems that our Australian colleagues have now “got their eye in” for complex thalloid liverwort Monocarpus sphaerocarpus – after many years…

In the Herbarium at RBGE, we store a huge number of sheets of archival quality paper with squashed and dried plant specimens stuck to them. These have been…

Spring Break’s a big thing in the US, and spring of 2005, Juan Carlos Villarreal and I spent ours on a road-trip down through Louisianna, looking for the…

Many new species are already included in natural history collections around the world, it’s just that nobody has yet got around to examining the material, recognising that it represents something…

Murphy’s threadwort (Telaranea murphyae) has had a singular position in the British flora. The species was described by renowned bryologist Jean Paton in 1965, from plants collected in…

Despite its internationally important bryophyte flora Scotland has relatively few truly endemic species (perhaps four), and even some of these have a rather ambiguous taxonomic status due to…

In conjunction with Dr Daniela Schill’s monographic work on Sphaerocarpos, we’ve been building a molecular phylogeny for the genus. We have attempted to extract DNA from 66 accessions,…

Not long ago, the only non-crop plant that the mainstream scientific community seemed to be aware of was the brassica Arabidopsis thaliana – easily cultivated, with a short…

Sitting in Edinburgh airport on a Monday morning, waiting for David Long to join me, checked in through to Trondheim via Copenhagen, I felt completely unprepared. The previous week…