Tag: Phylogeny
When conservation scientists are trying to decide which species are most in need of protection, the main consideration is usually how likely they are to become extinct, as…

I’m just back from field work in New Zealand with Yoan Coudert, a French CNRS funded researcher based at the Ecole Normale Supérieure in Lyon. A major objective…

As far as our 2013 RBGE MSc project proposal to generate a phylogeny of Octoblepharum goes, Juan Carlos Villarreal, Noris Salazar Allen and I were clearly not the…

One of the main problems with sampling largely from herbarium specimens, rather than from material that has been specifically collected for DNA work (rapidly dried in silica gel…

The genus Aitchisoniella contains a single species, A. himalayensis, which was described by Pakistani botanist Professor Shiv Ram Kashyap from plants that he collected in Mussoorie, Uttarakhand, India…

Sitting in Edinburgh airport on a Monday morning, waiting for David Long to join me, checked in through to Trondheim via Copenhagen, I felt completely unprepared. The previous week…
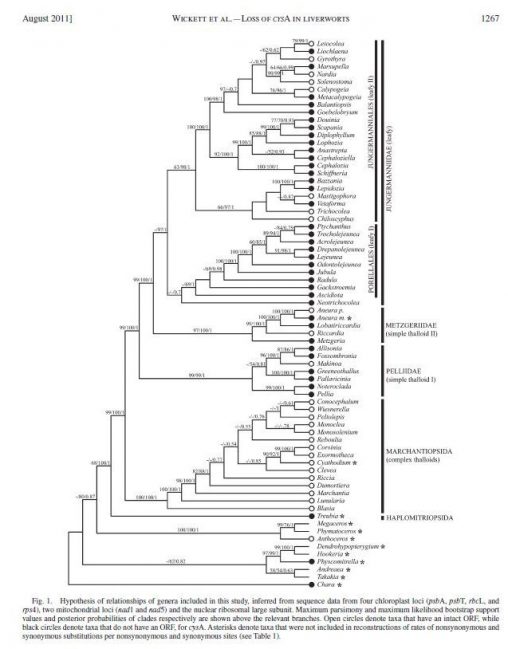
Although the exact relationships between the earliest land plant lineages are not yet well resolved, there is consensus that liverworts are one of the most ancient land plant…

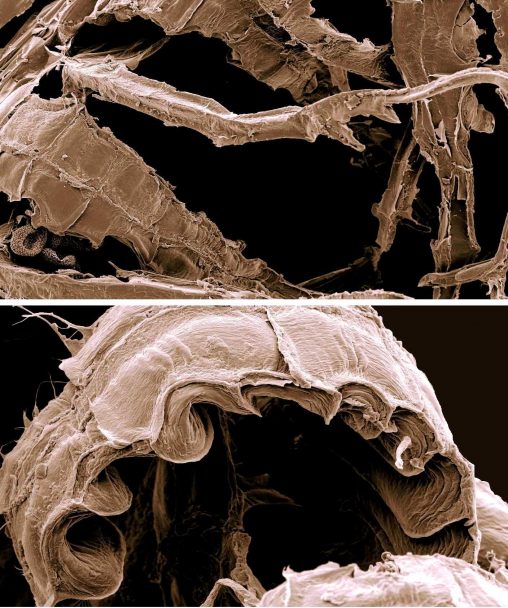
One of the earliest plastid genomes to be sequenced, in the late 1980s (Ohyama et al.), was that of Marchantia polymorpha, one of the commonest liverworts around town,…

Rather a while ago, back in 2003, we started working on a phylogeny of the complex thalloid liverworts at the Royal Botanic Garden Edinburgh (as a Molecular Phylogenetics…

As far as liverworts go, Monocarpus is a rather strange plant. It’s very small, in itself not that unusual for a bryophyte, but rather problematic if you need…

About 300 species of Inga (Leguminosae: Mimosoideae: Ingeae) grow in lowland and montane rain forest throughout the humid tropical zone, from Mexico to Uruguay. Most species diversity is…
Part 1: The Very Basics The analyses are finally over, you can fill in those blanks in the results section, and really start dealing with all those hypotheses…