Author: Neil Bell

Geography, and particularly climate, have distinguished the extreme western parts of Scotland from the rest of the country for thousands of years. Many of our rarest plant species…
When conservation scientists are trying to decide which species are most in need of protection, the main consideration is usually how likely they are to become extinct, as…

I’m just back from field work in New Zealand with Yoan Coudert, a French CNRS funded researcher based at the Ecole Normale Supérieure in Lyon. A major objective…

Despite its internationally important bryophyte flora Scotland has relatively few truly endemic species (perhaps four), and even some of these have a rather ambiguous taxonomic status due to…

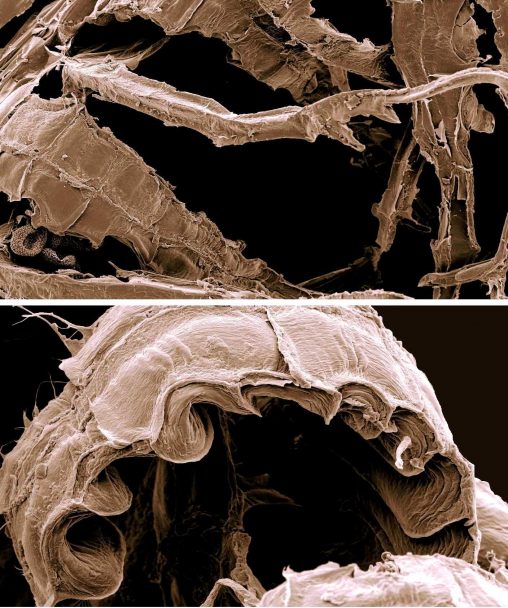
Colura calyptrifolia (or to give it its appropriately creepy-sounding common name, the Fingered Cowlwort), is one of our most fascinating UK liverworts. Absolutely tiny (the leaves are about…
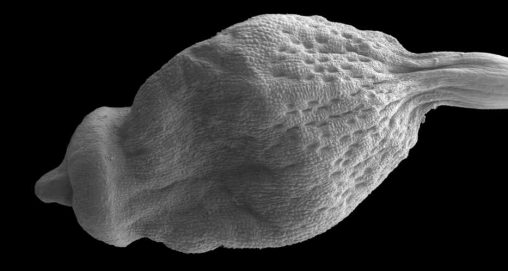
The relative structural simplicity of some groups of mosses can disguise their uniqueness, especially when simplified features have evolved multiple times within the same family from ancestors with…

What sorts of features provide the best clues about whether or not two plants are closely related? Sometimes it’s obvious – most people can correctly recognise a daffodil…