The tiny tricolour axillary hairs of the moss Leptobryum wilsonii subtly advertise its true relationships
What sorts of features provide the best clues about whether or not two plants are closely related? Sometimes it’s obvious – most people can correctly recognise a daffodil (Narcissus) by its general appearance, even if individual species vary a lot in shape, size and colour. That’s because the combination of six coloured sepals and a trumpet-like corona is pretty much unique to daffodils, and all daffodils are like this because they are closely related. Often it’s less obvious however. Non-botanists might think that what we call the sycamore in the UK (Acer pseudoplatanus) must be closely related to the London plane (Platanus × acerifolia) because of its very similarly shaped leaves. In fact it isn’t at all, and both trees are much closer to other species that have completely different leaves. Leaf shape in trees is often not a very good clue to relationship, because the same forms have evolved independently many times in different groups.

Leaf cells of Leptobryum wilsonii. Size, shape and other properties of leaf cells provide important characters for identifying mosses
It’s the same with mosses of course, except that the features we have to rely on are fewer, smaller, and often less complex. A couple of weeks ago, an American colleague sent me a packet containing a few stems of a moss he had collected on a remote island south of Tierra del Fuego. “I think this could be a new genus of Rhizogoniaceae”, he said. I had studied this family for my PhD, and certainly there was some undefinable gestalt quality this moss had that reminded me of these plants. Also the cell shape, while not typical for the family, was very close to that of one particular species in the group. However, it also looked rather like a Pohlia, a rather non-descript genus with spearhead-shaped leaves. Unfortunately this plant didn’t have many truly distinctive features – except one.
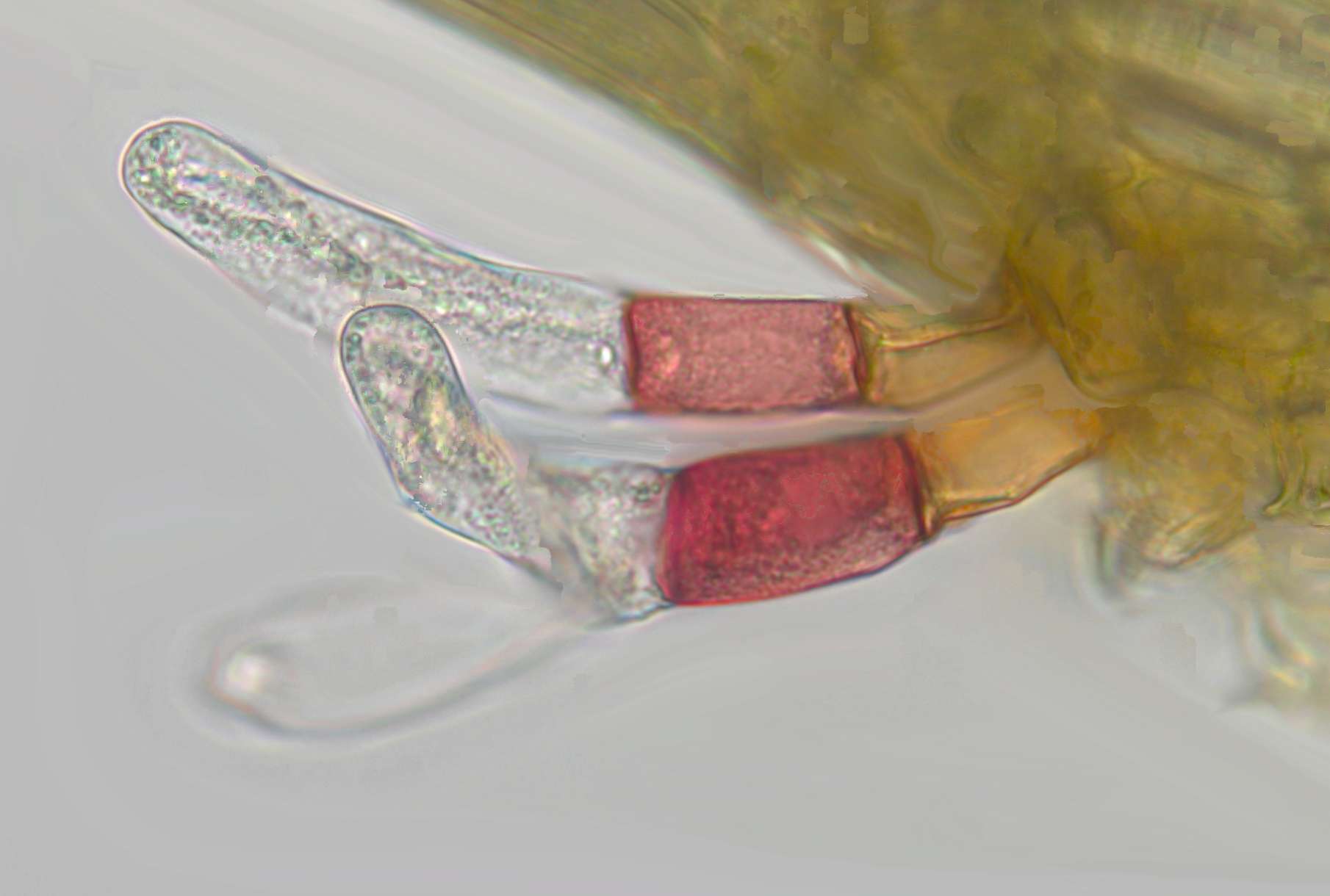
The weirdest thing about this moss was its axillary hairs. Axillary hairs are minute structures produced next to the leaves of mosses. They are just a few cells long and often only visible on the youngest parts of moss stems after the leaves have been removed. It’s thought that they may have a function in protecting the immature leaves and shoots when they are themselves just collections of a few cells. As the fully mature leaves of this moss are only a little over a millimetre long, you can imagine how small the axillary hairs are! Yet to a bryological eye, these particular axillary hairs are as ostentatious as a peacock’s tail feathers.

Axillary hairs are found in most mosses, just above the leaf insertion (i.e. in the leaf axil). They are composed of just a few cells and may secrete mucilage to protect the delicate tips of immature leaves or shoots
They had an unusual “tricolour” appearance with a small orange basal cell, surmounted by a larger bright pinkish-red cell, and then a really long, club-shaped transparent cell. What’s more, they were prominent and persistent, not falling off on the older parts of the stem as is often the case. While it was the brightly and differently coloured basal cells that really drew our attention to these structures, the single long, club-shaped top cell was the biggest clue to relationships.
This axillary hair morphology pointed towards a family called the Meesiaceae, a small group that includes the very common moss Leptobryum pyriforme, a species with very long, narrow leaves that is often found as a weed in glasshouses and in flowerpots. Our moss had completely different leaves – shorter, broader and spearhead-shaped. Nonetheless it transpired that it was Leptobryum wilsonii, now thought to be the only other species of Leptobryum, much less common than Leptobryum pyriforme and restricted to South America, South Africa and Antarctica. It was not an undescribed Rhizogoniaceae, as some aspects of its habit implied, nor a Pohlia, as the leaf form suggested. However, Leptobryum wilsonii is so unlike Leptobryum pyriforme superficially, and so much more like the unrelated Pohlia, that as recently as 2008 it was named Pohlia wilsonii and is still treated under that alternativename by some bryologists (in spite of its axillary hairs!).

Misleadingly, the spearhead-shaped leaves of Leptobryum wilsonii are very similar to those of the genus Pohlia
So when some features point towards one relationship and others towards a completely different affinity, how do we decide where the truth lies? The answer is like a more analytical version of how we tell if people are telling truth – if a particular feature is consistent with all the other information we have available and with the deep structure of that information, then we tend to have more confidence in it. However, if the consequences are that we would need to assume that a lot of other seemingly improbable things are also true (as when someone tells us that they have seen a ghost), then we are more sceptical. Sometimes we also simply trust some morphological characters more than others, just as we might trust some people more than others. We know that some features (such as leaf shape) often “lie” because they are prone to convergence in evolution, and so are more wary of them as guides to relationship.
The more features we have available as potential clues to relationship the better, because it makes it more likely that we will find a convincing consensus among the contradictory babble. That’s one reason why information from DNA sequences is now so important in taxonomy. It can provide a large enough quantity of unambiguous characters for us to find a statistically convincing consensus about one possible relationship over another.
As it happens, just recently some Japanese researchers were puzzled by a moss found submerged in lakes in coastal Antarctica. This moss was one of two species forming bizarre tower-like structures called “moss pillars” on the bottoms of these lakes.

“Moss pillars” in a lake in coastal East Anarctica (photograph courtesy of Satoshi Imura). The dominant species was identified as an aquatic form of Leptobryum wilsonii.
Like many mosses that occur as aquatics as well as on land, it had a strongly modified aquatic growth form that made it particularly difficult to identify. So they obtained DNA sequences from a number of samples, and were able to demonstrate that their aquatic plant was a form of Leptobryum wilsonii (Nakai et al. 2012, Kato et al. 2013). Furthermore, this data was also able to show very convincingly that Leptobryum wilsonii (whether in its aquatic or terrestrial forms) is indeed closely related to the familiar and very different Leptobryum pyriforme, and not at all to Pohlia. So leaf shape and the general habit of the plant are red herrings – features that have evolved independently in distantly related groups. For the plant we were attempting to identify those distinctive axillary hairs, despite their relative inconspicuousness, had been the best visible clue to its true relationships all along, pointing towards the surprising and incongruous affinity between the scruffy mosses in flower pots and the ones forming those unique “moss pillar” ecosystems in Antarctic lakes!
Nakai, R., Abe, T., Baba, T., Imura, S., Kagoshima, H., Kanda, H., Kohara, Y., Koi, A., Niki, H., Yanagihara, K., Naganuma, T. 2012. Eukaryotic phylotypes in aquatic moss pillars inhabiting a freshwater lake in East Antarctica, based on 18S rRNA gene analysis. Polar Biol. 35: 1495-1504.
Kato, K., Arikawa, T., Imura, S., Kanda, H. 2013. Molecular identification and phylogeny of an aquatic moss species in Antarctic lakes. Polar Biol. 36: 1557-1568.