One of the most recognisable groups in the bryophytes, the complex thalloid liverwort genus Marchantia, has just become a bit larger. We have sunk Preissia and Bucegia into it, because in molecular phylogenies they are both phylogenetically nested within Marchantia (Villarreal et al. 2015). Although this only adds two species to the genus (Preissia quadrata and Bucegia romanica), taking the total number of species recognised from 36 to 38, it also broadens the range of plant morphologies that occurs in Marchantia.

Maximum likelihood topology for Marchantia adapted from Villarreal et al. 2015; * indicates bootstrap support of 100% and posterior probabilites of 1
Preissia quadrata (Scopoli) Nees returns to an earlier name, Marchantia quadrata Scopoli. This is the name under which it was first described, by Scopoli, in 1772. For Bucegia romanica Rad., first described by Radian (1903), a new combination has been made, Marchantia romanica (Rad.) Long, Crandall-Stotler, Forrest & Villarreal.
A new subgenus, Marchantia subg. Preissia, has been created for these species. A third species that was collected in China by David Long, and was included in the molecular phylogeny, has not yet been named, but will also belong to subgenus Preissia. Morphologically, the new species is most similar to Marchantia quadrata.
 Marchantia romanica is known from northern Europe and northern North America (e.g. Evans 1917, Konstantinova et al. 2014). Photographs of fertile plants, and transverse sections showing 2-3 layers of filament-less air chambers, can be seen in Konstantinova et al.’s report. The hollow air chambers they illustrate are one of the clearest features that separated Bucegia from Marchantia and Preissia.
Marchantia romanica is known from northern Europe and northern North America (e.g. Evans 1917, Konstantinova et al. 2014). Photographs of fertile plants, and transverse sections showing 2-3 layers of filament-less air chambers, can be seen in Konstantinova et al.’s report. The hollow air chambers they illustrate are one of the clearest features that separated Bucegia from Marchantia and Preissia.

Marchantia quadrata carpocephalum, photographed by Des Callaghan
Marchantia quadrata is also a northern hemisphere species; it is widespread in the British Isles, and a map and photographs are available in the British Bryological Society Field Guide. Like Marchantia romanica, this species lacks gemmae cups. Its two ranks of ventral scales could also be used to separate Preissia from Marchantia, which typically has 4 or more rows of ventral scales. Two subspecies are recognised in Schuster (1992), one more southerly and dioicous (ssp quadrata) and the other northern and monoecious (ssp hyperborea) (Schuster 1985). From our data, using DNA regions from genes that were chosen to show relationships between genera rather than within species, genetic differences can be seen between a collection from the South Ebudes off the west coast of Scotland, and one from Snowbird, Utah in the United States. Molecular sampling from across the range of Marchantia quadrata may reveal a complex of several species. Indeed, this was anticipated by Schuster (1992, p. 366), who described the range of morphological variation within the species as “astonishing”.
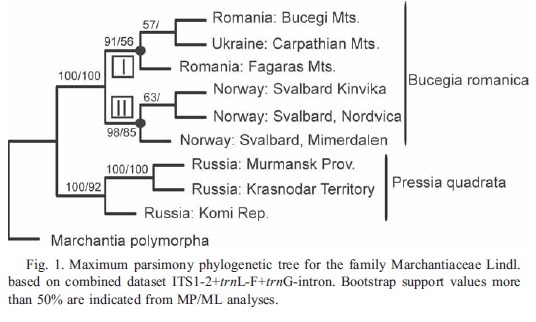
extracted from Konstantinova et al. 2014: phylogenetic variation between accessions of Bucegia and Preissia
Konstantinova et al. (2014) generated DNA sequences for the nuclear ITS region, and two plastid regions, trnL-F and trnG, for six accessions of Marchantia romanica (“Bucegia”, from Romania, Ukraine and Svalbard), and three accessions of Marchantia quadrata (“Pressia”, from Russia). Unfortunately the regions that they sequenced do not overlap with the sequencing in our study, so we cannot combine the datasets. However, they do rather intriguingly resolve two clades within Marchantia romanica (“Bucegia”), one from mainland Europe, and the other from Svalbard. Again, molecular sampling from across the range of this taxon could reveal several distinct lineages. Adding samples from North America seems a priority.
Thus, although we have only added two species to Marchantia, the number of species is likely to rise with additional molecular sampling, in line with a clearly necessary taxonomic revision of Marchantia subgenus Preissia.
David G. Long, Laura L. Forrest, Juan Carlos Villarreal & Barbara J Crandall-Stotler. 2016. Taxonomic changes in Marchantiaceae, Corsiniaceae and Cleveaceae (Marchantiidae, Marchantiophyta). Phytotaxa 252 (1): 077-080.

Gary Powell
Very interesting !
Can you try to describe to me what on earth – ” bootstrap support of 100% and posterior probabities of 1″ – means ?
I wonder how many other Bryophytes will phylogenetically combine (or be separated )
?
Amateur bryologist ( been studying them for 4 yrs.)
Laura Forrest
Gary, bootstrapping is a data resampling method, and posterior probabilites relate to the likelihood of a particular relationship – they’re both indications of the strength of support in the data for the relationships that are shown. 100% bootstrap support and posterior probability of 1 are the highest values, indicating strong support.