Tag: BryophytePage 1 of 3
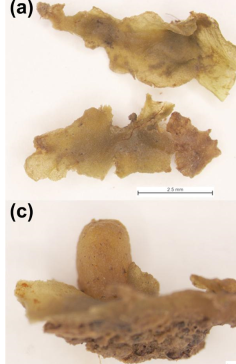
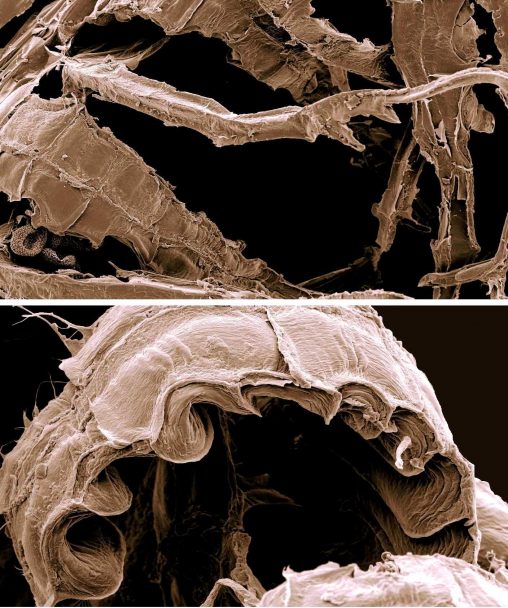
We have chosen a lectoype for Riccardia fuscovirens Lindb. (i.e., a type specimen that is chosen after a name has been published, but from the original specimens or illustrations that the author of the name would have been influenced by when they were first recognizing the taxon)
When conservation scientists are trying to decide which species are most in need of protection, the main consideration is usually how likely they are to become extinct, as…
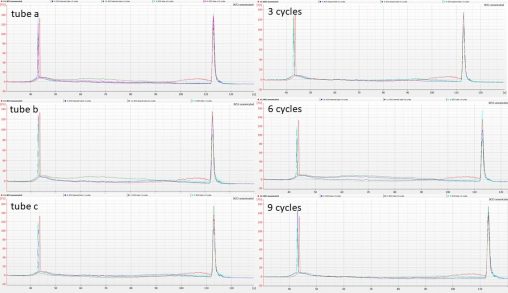
Following on from the rather unpredictable results we obtained from fragmenting duplicate aliquots of CTAB-extracted Polytrichum DNA in the Bioruptor, Isuru cleaned aliquots of IK31 and IK53 using…
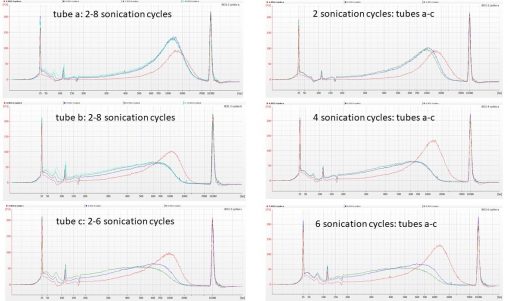
We started our lab work on the Polytrichum hybrid baits project on the 1st of October, by normalising some CTAB-extracted DNA with 0.1X TE to 55 µL of…

The current Next Gen Sequencing lab project at the Botanics involves looking at the phylogeny of Polytrichum section Polytrichum, using hybrid capture. The work will form part of…

The leafy liverwort genus Nardia has five species that are known to occur in Scotland. Of these five species that occur in Scotland, four are described in Smith’s…

Dr Des Callaghan spends rather a lot of his time chasing after rare things. He’s an environmental consultant with many strings to his bow, but a particular specialisation…

Life gets littered with untold stories; here’s one that did get told, briefly, and then got forgotten. It was told at the Botany meeting in Austin in August…

Four years ago, we took an overnight trip to one of the four gardens of the Royal Botanic Garden Edinburgh: Benmore, in Argyll (Sunday 2nd – Monday 3rd February…

The Shawnee National Forest skirts the midwestern town of Carbondale, which is home to one of the campuses of Southern Illinois University. It’s also one of the prettiest…

One of North America’s endemic hornworts, Phaeoceros proskaueri Stotler, Crand.-Stotl. & W.T.Doyle [also known as Paraphymatoceros proskaueri (Stotler, Crand.-Stotl. & W.T.Doyle) J.C.Villarreal & Cargill] was described from plants collected in the Monterey Bay…

During a family holiday to Santiago, Panama in June/July 2011, we snuck in a short bryologising trip, first heading west along the Pan-American Highway, then north, to the…

The Botany 2004 meeting was in Snowbird, Utah – a chance to see a different part of the United States (and, of course, to present our research to…

In April 2004, I flew north from Illinois to met up with a botanical friend, Dr Zoe Badcock. Our meeting point was Vancouver, British Columbia; from there we…

There’s something quite melancholy about going back through all the little paper packets of voucher specimens, remembering who and where you were when you collected them, and thinking…

The Science building at the Botanics closes down between Christmas and New Year, so any last bits of work for the year have to be packed up and…

Now that we have six wild-collected accessions of Plagiochasma currently growing on public display in the RBGE Arid House, from China, the US (Texas) and Saudi Arabia, I’ve…

I’m just back from field work in New Zealand with Yoan Coudert, a French CNRS funded researcher based at the Ecole Normale Supérieure in Lyon. A major objective…

After RBGE’s initial involvement in land plant DNA barcode marker selection, culminating in a couple of 2009 papers that both utilized bryophyte barcoding data sets, we started a…

Enigmatic and isolated although it is, it seems that our Australian colleagues have now “got their eye in” for complex thalloid liverwort Monocarpus sphaerocarpus – after many years…